 www.PDImages.com")
Few people can have escaped the newspaper headlines over the past few months. The fertility doctor Panos Zavos claimed to have cloned a human baby, while a team of researchers in South Korea also presented their clones to the world. So what's the difference between these stories? And are we really going to see cloned babies peering out from their prams in the near future? In this first article we will spy on the earliest moments of life: how an egg and sperm make a ball of cells that eventually become a baby. Then we can ask if, and how, these processes can be bypassed by cloning. The first thing to understand is how babies are made - no sniggering at the back, please! To do this, we need to know a bit about eggs and sperm. Eggs are relatively large cells and contain lots of biological goodies required for early development. They are also rather unusual in that they are frozen in the act of dividing their DNA from a full set of chromosomes to half a set (see figure 1, below). This division is important because the sperm also carries half a set: together they make up a full set in the new baby.
At the moment of fertilisation the sperm enters the egg, and things start getting exciting (at least, in biological terms). The following descriptions show what happens in mouse development: we currently do not know whether human development is exactly the same, but it is likely that many events are similar. When the sperm goes in, it reactivates the division process in the egg, causing the arrested chromosome half-sets to separate. The unwanted chromosomes are booted out and form a little cell called the polar body, with no further part to play in the unfolding developmental drama (see figure 1, below). The remaining egg chromosomes organise into a ball-like structure termed a pronucleus. At the same time, proteins in the egg begin to unpack the sperm DNA, expanding it to form another pronucleus (see figure 1, below).
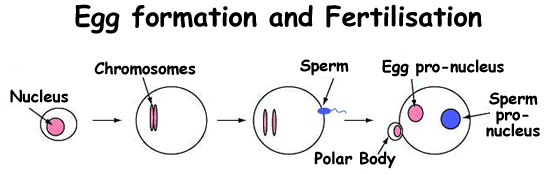 | |||||||
1. An early egg cell. Humans carry 23 pairs of chromo-somes. We inherit one half of each pair from our mothers, and the other half from our fathers. These half-sets of chromosomes are produced in a special form of cell division called meiosis which produces eggs and sperm. | 2. A mature egg. In this egg, for simplicity, only one pair of chromosomes is shown (in pink). The process of cell division remains frozen until the egg is fertilised by a sperm | 3. Fertilisation. When a sperm penetrates the egg it kick starts the completion of meiosis. One of each pair of the egg's chromosomes is randomly ejected from the cell to form a structure called a polar body. The remaining chromosomes form the egg pro-nucleus. | 4. One cell embryo. The half-sets of chromosomes from the sperm are unpacked to form the sperm pro-nucleus. As each pro-nucleus contains half the normal chromosome number, together they produce a cell with a full complement of genetic material. | ||||
Figure 1 - The formation of a mature egg and a single celled embryo. | |||||||
But strange things happen to the sperm DNA: it is stripped of methylation, a special molecular tag that helps the cell to use its genes properly. In the first few hours of development other dynamic changes happen to the sperm pronucleus, especially alterations in the DNA packing proteins within it. These changes in methylation and packing proteins are termed "epigenetic modifications", as they affect the DNA without actually changing the underlying DNA sequence - only the tags and markers around it. Epigenetic marks are important because they tell the cell which genes to use in different types of cells. This is crucial because all our cells contain essentially the same DNA (and therefore the same genes): it is the different patterns of gene usage that give all our cell types their distinct characteristics. Some epigenetic marks act as silencing signals for genes, while some have an activating effect. For example, in a liver cell, liver-type genes would have activating marks while muscle genes would have silencing marks. Conversely, in a muscle cell the muscle-specific genes have activating tags whereas the liver genes are silenced.
Many epigenetic changes take place while the embryo is still a single cell, and these mainly occur within the sperm pronucleus. These molecular upheavals are essential for reprogramming the sperm, so it can be used correctly in the next steps of development. But the epigenetic adventures don't stop at the one-cell stage. The new embryo divides into two cells, then four, then eight and so on until it is a ball of around a hundred cells (see figure 2). Throughout this flurry of activity, more molecular tags are removed from the DNA while other epigenetic modification patterns are established. Eventually, after 4 to 5 days, the ball of cells begins to take shape. A cavity forms and fills with fluid, pushing the cells outwards until the ball is almost hollow and looks rather like a football. We call this a "blastocyst". But the blastocyst is not quite hollow, because lurking on one side is a small clump of cells, somewhat obviously named the "inner cell mass". It is from this unpromising cluster that we all grew: these are the stem cells of the embryo. It is also clear at this stage that there are distinct epigenetic differences between the outer cells and the inner cell mass.
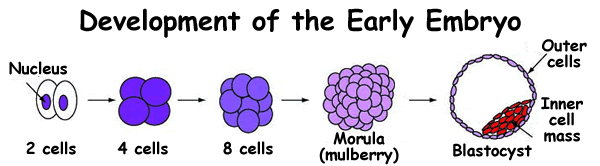 | |||||
| The 2 pronuclei merge, producing a cell with a complete set of 46 chromosomes (23 pairs), The cell begins to divide, giving rise to 2, then 4, then 8 cells, and so on. | The cells continue to divide, undergoing further epi-genetic changes until they eventually produce a cluster of cells resembling a mulberry, called a morula. | A blastocyst is a hollow cell ball containing a cluster of stem cells (the inner cell mass). The inner cell mass (which gives rise to the baby itself) is epi-genetically distinct from the outer cells, which produce the placenta. | |||
Figure 2 - The development of the early embryo | |||||
By this time, the embryo has made it to the uterus (womb). The outer cells of our embryonic football start invading the inner wall of the uterus and eventually make the placenta, the large organ that allows nutrients and waste to be exchanged between the mother and the baby. Meanwhile, the small clump of stem cells start dividing and undergo yet more epigenetic changes. An amazing programme of gene activity starts in these modest cells, causing them to organise and create all the parts of a new life. In the next article we'll find out how how cloning fits in to this natural developmental agenda.










Comments
Add a comment